| Reactivity | HuSpecies Glossary |
| Applications | ELISA |
| Conjugate | Biotin |
| Source | N/A |
| Assay Type | Solid Phase Sandwich ELISA |
| Inter-Assay | See PDF Datasheet for details |
| Intra-Assay | See PDF Datasheet for details |
| Spike Recovery | See PDF Datasheet for details |
| Sample Volume | See PDF Datasheet for details |
| Gene | CRP |
| Dilutions |
|
|
| Application Notes | No significant interference observed with available related molecules. |
|
| Publications |
|
| Storage | Store the unopened product at 2 - 8 °C. Do not use past expiration date. |
![Immunohistochemistry Albumin Antibody (188835) [Unconjugated] - Serum](https://images.novusbio.com/images/antibody/Albumin_MAB1455_Immunohistochemistry_23317.jpg)
![Immunocytochemistry Albumin Antibody (188835) [Unconjugated] - Serum](https://images.novusbio.com/images/antibody/Albumin_MAB1455_Immunocytochemistry__Immunofluorescence_17777.jpg)
![Intracellular Staining by Flow Cytometry Albumin Antibody (188835) [Unconjugated] - Serum](https://images.novusbio.com/images/antibody/Albumin_MAB1455_Flow_Cytometry_15110.jpg)







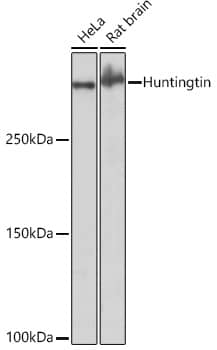




| Gene Symbol | CRP |



![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/DATA_IL6_M6000_ELISA_936.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/IL-6_M6000_ELISA_415.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/m6000b_mouse-il-6-quantikine-elisa-kit-1752025024034.jpg)
![SDS-Page TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNF-alpha_210-TA_256.jpg)
![Bioactivity TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNFalpha_210TA_1658.jpg)
![SEC-MALS TNF-alpha [Unconjugated]](https://images.novusbio.com/images/210-ta_recombinant-human-tnf-alpha-protein-sec-mals-35202312244..jpg)
![Immunohistochemistry Insulin Antibody (182410) [Unconjugated]](https://images.novusbio.com/images/antibody/mab1417_human-bovine-mouse-insulin-mab-clone-182410-immunohistochemistry-308202115145.jpg)
![Immunocytochemistry Insulin Antibody (182410) [Unconjugated]](https://images.novusbio.com/images/antibody/Insulin_MAB1417_Immunocytochemistry_9376.jpg)
![N/A Adiponectin/Acrp30 [HRP]](https://images.novusbio.com/images/elisa/DATA_Adiponectin_DRP300_ELISA_812.jpg)
![N/A Adiponectin/Acrp30 [HRP]](https://images.novusbio.com/images/elisa/Adiponectin_DRP300_ELISA_173.jpg)
![N/A Adiponectin/Acrp30 [HRP]](https://images.novusbio.com/images/elisa/DATA_Adiponectin_DRP300_ELISA_813.jpg)
![N/A IL-10 [Biotin]](https://images.novusbio.com/images/elisa/DATA_IL10_DY417_ELISA_2014.jpg)
![N/A Leptin/OB [HRP]](https://images.novusbio.com/images/elisa/Leptin_DLP00_ELISA_134.jpg)
![N/A Leptin/OB [HRP]](https://images.novusbio.com/images/elisa/DATA_Leptin_DLP00_ELISA_749.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/CCL2_DCP00_ELISA_67.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/DATA_CCL2_DCP00_ELISA_633.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/DATA_CCL2_DCP00_ELISA_634.jpg)
 - BSA Free")
![Bioactivity IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/protein/IL-1_beta_201-LB_306.jpg)
![SDS-Page IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/protein/IL-1_beta_201-LB_304.jpg)
![SEC-MALS IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/201-lb_recombinant-human-il-1-beta-il-1f2-protein-sec-mals-123202491556.jpg)
![Bioactivity IL-2 [Unconjugated]](https://images.novusbio.com/images/202-il_recombinant-human-il-2-protein-bioactivity-174202314946.jpg)
![N/A Pentraxin 2/SAP [HRP]](https://images.novusbio.com/images/elisa/DATA_Pentraxin_2_MPTX20_ELISA_1018.jpg)
![N/A Pentraxin 2/SAP [HRP]](https://images.novusbio.com/images/elisa/Pentraxin_2_MPTX20_ELISA_310.jpg)