| Applications | Binding Activity |
| Details of Functionality | Measured by its binding ability in a functional ELISA. When Mannan is coated at 0.2
μg/mL (100 μL/well), the concentration of Recombinant Mouse MMR/CD206
that produces 50% of the optimal binding response is
approximately 0.1-0.6 μg/mL. |
| Source | Mouse myeloma cell line, NS0-derived mouse MMR/CD206 protein Leu19-Ala1388, with a C-terminal 6-His tag |
| Accession # | |
| N-terminal Sequence | Leu19 |
| Protein/Peptide Type | Recombinant Proteins |
| Gene | Mrc1 |
| Purity | >90%, by SDS-PAGE visualized with Silver Staining and quantitative densitometry by Coomassie® Blue Staining |
| Endotoxin Note | <0.10 EU per 1 μg of the protein by the LAL method. |
| Dilutions |
|
|
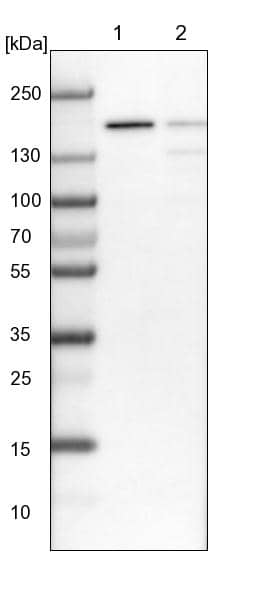
| Theoretical MW | 156 kDa (monomer). Disclaimer note: The observed molecular weight of the protein may vary from the listed predicted molecular weight due to post translational modifications, post translation cleavages, relative charges, and other experimental factors. |
|
| SDS-PAGE | 141-170 kDa, reducing conditions |
|
| Publications |
|
| Storage | Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
|
| Buffer | Lyophilized from a 0.2 μm filtered solution in PBS with BSA as a carrier protein. |
| Purity | >90%, by SDS-PAGE visualized with Silver Staining and quantitative densitometry by Coomassie® Blue Staining |
| Reconstitution Instructions | Reconstitute at 100 μg/mL in sterile PBS. |




































The concentration calculator allows you to quickly calculate the volume, mass or concentration of your vial. Simply enter your mass, volume, or concentration values for your reagent and the calculator will determine the rest.




![N/A IL-10 [Biotin]](https://images.novusbio.com/images/elisa/DATA_IL10_DY417_ELISA_2014.jpg)
![SDS-Page TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNF-alpha_210-TA_256.jpg)
![Bioactivity TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNFalpha_210TA_1658.jpg)
![SEC-MALS TNF-alpha [Unconjugated]](https://images.novusbio.com/images/210-ta_recombinant-human-tnf-alpha-protein-sec-mals-35202312244..jpg)
![Bioactivity IL-4 [Unconjugated]](https://images.novusbio.com/images/protein/6507-ilcf_recombinant-human-il-4-cho-expressed-protein-cf-bioactivity-272020133214.jpg)
![Bioactivity CTLA-4 [Unconjugated]](https://images.novusbio.com/images/protein/CTLA4_7268CT_2293.jpg)
, LYVE1 (b), and MRC1 (c) in the meninges of human post mortem brain showing no signs of neuropathology. These images are taken from a 38 year old male (sample P17/07, Table 1), and confirmed in n = 2 additional samples. P parenchyma. Scale = 150 µm (a); 40 µm (b); and 20 µm (c). d–f DAB-IHC with single antibodies detects VEGFR3 (b), LYVE1 (c), and MRC1 (d) in elderly human meninges (age: 89–92) with evidence of neuropathology and confirmed in n = 3 brains (Table 1). P, parenchyma. Scale = 20 µm. g–p IHC with fluorescent antibodies detects human meningeal cells that co-express MRC1 (h, m, yellow), LYVE1 (i, n, white), and VEGFR3 (j, o, green). Nuclei/RNA are labelled with DAPI (g, l, blue) and images are merged in (k, p). Scale = 10 µm Image collected and cropped by CiteAb from the following publication (https://pubmed.ncbi.nlm.nih.gov/31696318), licensed under a CC-BY license. Not internally tested by R&D Systems.")