 972-EMN) binds Recombinant SARS-CoV-2 Spike RBD Fc Chimera (10499-CV) in a functional ELISA." title="Recombinant Human EMMPRIN/CD147 Fc Chimera (972-EMN) binds Recombinant SARS-CoV-2 Spike RBD Fc Chimera (10499-CV) in a functional ELISA." />
972-EMN) binds Recombinant SARS-CoV-2 Spike RBD Fc Chimera (10499-CV) in a functional ELISA." title="Recombinant Human EMMPRIN/CD147 Fc Chimera (972-EMN) binds Recombinant SARS-CoV-2 Spike RBD Fc Chimera (10499-CV) in a functional ELISA." />
| Reactivity | HuSpecies Glossary |
| Applications | Bioactivity |
| Format | Carrier-Free |
| Additional Information | Isoform 2. aa 22-205 |
||||||||
| Details of Functionality | Measured by the ability of the immobilized protein to induce active MMP-1 secretion by NHLF human normal lung fibroblasts. The ED50 for this effect is 2‑8 µg/mL. Measured by its binding ability in a functional ELISA with Recombinant
SARS-CoV-2 Spike RBD Fc Chimera
(Catalog #
10499-CV). |
||||||||
| Source | Mouse myeloma cell line, NS0-derived human EMMPRIN/CD147 protein
|
||||||||
| Accession # | |||||||||
| N-terminal Sequence | Thr25 |
||||||||
| Structure / Form | Disulfide-linked homodimer |
||||||||
| Protein/Peptide Type | Recombinant Proteins |
||||||||
| Gene | BSG |
||||||||
| Purity | >95%, by SDS-PAGE under reducing conditions and visualized by silver stain |
||||||||
| Endotoxin Note | <0.10 EU per 1 μg of the protein by the LAL method. |
| Dilutions |
|
|
| Theoretical MW | 47.4 kDa (monomer). Disclaimer note: The observed molecular weight of the protein may vary from the listed predicted molecular weight due to post translational modifications, post translation cleavages, relative charges, and other experimental factors. |
|
| SDS-PAGE | 60 - 65 kDa, under reducing conditions. |
|
| Publications |
|
| Storage | Use a manual defrost freezer and avoid repeated freeze-thaw cycles.
|
| Buffer | Lyophilized from a 0.2 μm filtered solution in PBS with Trehalose. |
| Purity | >95%, by SDS-PAGE under reducing conditions and visualized by silver stain |
| Reconstitution Instructions | Reconstitute at 100 μg/mL in sterile PBS. |















 Protein, CF")
The concentration calculator allows you to quickly calculate the volume, mass or concentration of your vial. Simply enter your mass, volume, or concentration values for your reagent and the calculator will determine the rest.
| Gene Symbol | BSG |
| Uniprot |


![Bioactivity CTLA-4 [Unconjugated]](https://images.novusbio.com/images/protein/CTLA4_7268CT_2293.jpg)
![N/A MMP-2 [HRP]](https://images.novusbio.com/images/elisa/DATA_MMP2_MMP200_ELISA_1003.jpg)
![N/A MMP-2 [HRP]](https://images.novusbio.com/images/elisa/MMP2_MMP200_ELISA_474.jpg)
![N/A MMP-9 [HRP]](https://images.novusbio.com/images/elisa/DATA_MMP9_DMP900_ELISA_770.jpg)
![N/A MMP-9 [HRP]](https://images.novusbio.com/images/elisa/MMP-9_DMP900_ELISA_148.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/VEGF_DVE00_ELISA_208.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/DATA_VEGF_DVE00_ELISA_871.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/DATA_VEGF_DVE00_ELISA_872.jpg)
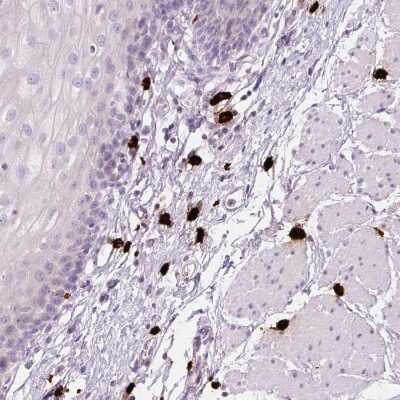
![Immunohistochemistry MMP-1 Antibody (36665) [Unconjugated]](https://images.novusbio.com/images/antibody/MMP-1_MAB901_Immunohistochemistry_9677.jpg)
![Western Blot MMP-1 Antibody (36665) [Unconjugated]](https://images.novusbio.com/images/antibody/MMP-1_MAB901_Western_Blot_9939.jpg)

![Western Blot ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Western_Blot_5097.jpg)
![Knockout Validated ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Knockout_Validated_22864.jpg)
![Immunohistochemistry ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Immunohistochemistry_20696.jpg)

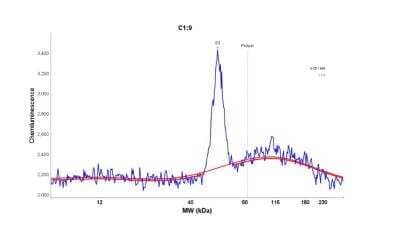



 followed by HRP-conjugated Anti-Goat IgG Secondary Antibody (Catalog # HAF019). A specific band was detected for EMMPRIN/CD147 at approximately 38-50 kDa (as indicated). This experiment was conducted under reducing conditions and using Immunoblot Buffer Group 1.")
{kind=link}