| Reactivity | HuSpecies Glossary |
| Applications | ELISA |
| Conjugate | HRP |
| Background | The QuantiGlo Human Endothelin-1 Chemiluminescent Immunoassay is a 4.5 hour solid phase ELISA designed to measure human Endothelin-1 levels in cell culture supernates, serum, EDTA plasma, and urine without extraction. It contains synthetic human Endothelin-1 and antibodies raised against the synthetic factor. This immunoassay has been shown to accurately quantitate human Endothelin-1. Result...s obtained using natural human Endothelin-1 showed dose curves that were parallel to the standard curves obtained using the QuantiGlo kit standards. These results indicate that this kit can be used to determine relative mass values of natural human Endothelin-1. Show More |
| Specificity | Natural and synthetic human Endothelin-1 |
| Source | N/A |
| Inter-Assay | See PDF Datasheet for details |
| Intra-Assay | See PDF Datasheet for details |
| Spike Recovery | See PDF Datasheet for details |
| Sample Volume | See PDF Datasheet for details |
| Gene | EDN1 |
| Dilutions |
|
|
| Application Notes | No significant interference observed with available related molecules. |
|
| Publications |
|
| Storage | Store the unopened product at 2 - 8 °C. Do not use past expiration date. |



![Immunohistochemistry ACE/CD143 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ACE_AF1513_Immunohistochemistry_6703.jpg)
![Western Blot ACE/CD143 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ACE_AF1513_Flow_Cytometry_20028.jpg)
![Simple Western ACE/CD143 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ACE_AF1513_Simple_Western_20109.jpg)

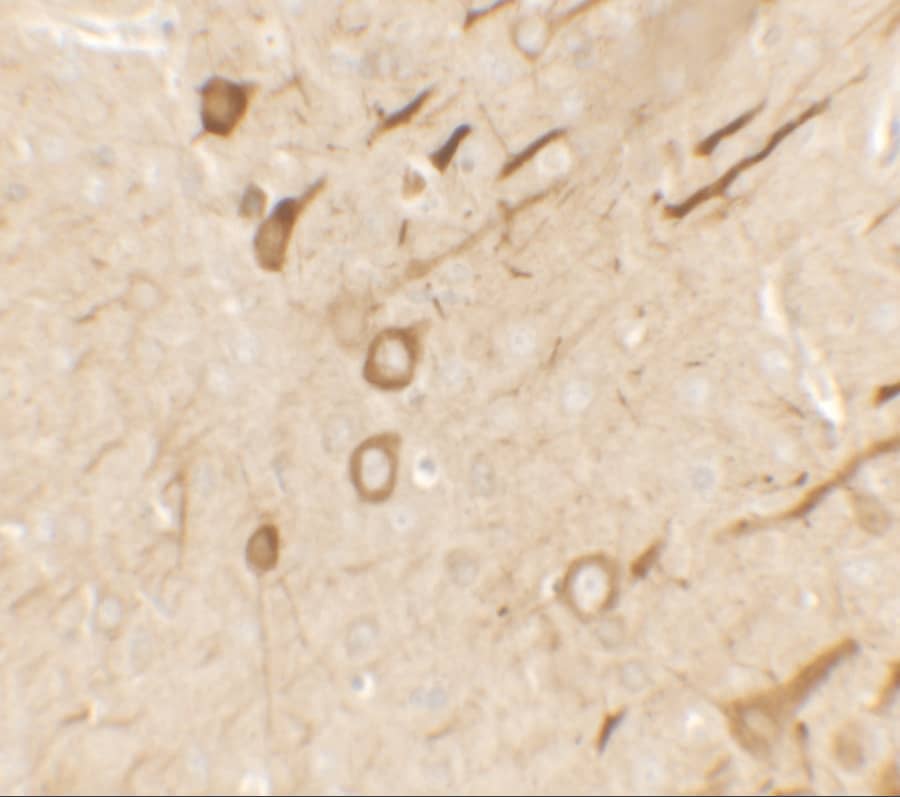

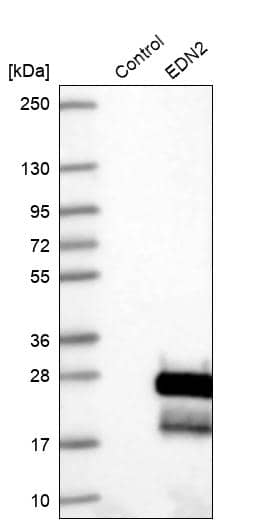
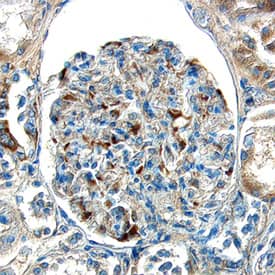
| Gene Symbol | EDN1 |
















![SDS-Page TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNF-alpha_210-TA_256.jpg)
![Bioactivity TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNFalpha_210TA_1658.jpg)
![SEC-MALS TNF-alpha [Unconjugated]](https://images.novusbio.com/images/210-ta_recombinant-human-tnf-alpha-protein-sec-mals-35202312244..jpg)


![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/DATA_IL6_M6000_ELISA_936.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/IL-6_M6000_ELISA_415.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/m6000b_mouse-il-6-quantikine-elisa-kit-1752025024034.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/VEGF_DVE00_ELISA_208.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/DATA_VEGF_DVE00_ELISA_871.jpg)
![N/A VEGF [HRP]](https://images.novusbio.com/images/elisa/DATA_VEGF_DVE00_ELISA_872.jpg)
![Western Blot ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Western_Blot_5097.jpg)
![Knockout Validated ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Knockout_Validated_22864.jpg)
![Immunohistochemistry ERK2 Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/ERK2_AF1230_Immunohistochemistry_20696.jpg)



![Immunohistochemistry Insulin Antibody (182410) [Unconjugated]](https://images.novusbio.com/images/antibody/mab1417_human-bovine-mouse-insulin-mab-clone-182410-immunohistochemistry-308202115145.jpg)
![Immunocytochemistry Insulin Antibody (182410) [Unconjugated]](https://images.novusbio.com/images/antibody/Insulin_MAB1417_Immunocytochemistry_9376.jpg)
![Immunocytochemistry/ Immunofluorescence Renin Antibody [Unconjugated] - 1](https://images.novusbio.com/images/af4277_mouse-renin-1-affinity-purified-polyclonal-ab-41202412394940.jpg)
![Immunohistochemistry Renin Antibody [Unconjugated] - 1](https://images.novusbio.com/images/antibody/Renin_AF4277_Immunohistochemistry_17351.jpg)
![ELISA Renin Antibody [Unconjugated] - 1](https://images.novusbio.com/images/af4277_mouse-renin-1-affinity-purified-polyclonal-ab-elisa-639201778625485467.jpg)
![Flow Cytometry: Endothelin-1 Antibody (TR.ET.48.5) [NB300-526] - Analysis of Endothelin 1 in 3T3 cells compared to an isotype control (blue).](https://images.novusbio.com/images/Endothelin-1-Antibody-TR-ET-48-5-Flow-Cytometry-NB300-526-img0012.jpg "Flow Cytometry: Endothelin-1 Antibody (TR.ET.48.5) [NB300-526] - Analysis of Endothelin 1 in 3T3 cells compared to an isotype control (blue).")