After August 17, 2026, Novus Biologicals products and services will no longer be available on this website; you will access all products and services on rndsystems.com. Create your R&D Systems online account today.
Recombinant Mouse Siglec-1 Fc Chimera Protein, CF Summary
Details of Functionality
Measured by the ability of the immobilized protein to support the adhesion of human red blood cells. Kelm, S. et al. (1994) Current Biology 4:965. The ED50 for this effect is 0.6-3 μg/mL.
Source
Mouse myeloma cell line, NS0-derived mouse Siglec-1/CD169 protein
>90%, by SDS-PAGE under reducing conditions and visualized by silver stain
Endotoxin Note
<0.01 EU per 1 μg of the protein by the LAL method.
Applications/Dilutions
Dilutions
Bioactivity
Theoretical MW
201.7 kDa (monomer). Disclaimer note: The observed molecular weight of the protein may vary from the listed predicted molecular weight due to post translational modifications, post translation cleavages, relative charges, and other experimental factors.
SDS-PAGE
190-205 kDa under reducing conditions
Publications
Read Publications using 5610-SL in the following applications:
Siglecs are sialic acid specific I-type lectins that belong to the immunoglobulin superfamily. Structurally, they are transmembrane proteins with an N-terminal Ig-like V-set domain followed by varying numbers of Ig-like C2-set domains (1, 2). Mouse Siglec-1, also known as sialoadhesin and CD169, is a 175 - 185 kDa glycoprotein that consists of a 1619 amino acid (aa) extracellular domain (ECD) with one Ig-like V-set domain and 16 Ig‑like C2-set domains, a 21 aa transmembrane segment, and a 35 aa cytoplasmic domain (3, 4). Within the ECD, mouse Siglec-1 shares 73% and 83% aa sequence identity with human and rat Siglec-1, respectively. Alternate splicing generates a soluble form of the ECD and a soluble isoform that is truncated following the first three Ig-like domains (3). Siglec-1 expression is restricted to lymph node and spleen macrophages and some tissue macrophages (4). The adhesive function of Siglec-1 is supported by the N-terminal Ig-like domain which shows a selectivity for alpha -2,3-linked sialic acid residues (4 - 6). Siglec-1 binds a number of sialylated molecules including the mannose receptor, MGL1, MUC1, PSGL-1, and different glycoforms of CD43 (7 - 10). Its binding capacity can be masked by endogenous sialylated molecules (11, 12). The sialylated and sulfated N-linked carbohydrates that modify Siglec-1 itself are required for ligand binding (7, 8). Siglec-1 is expressed on dendritic cells following rhinovirus exposure, and these DC promote T cell anergy (13). It is also induced on circulating monocytes during systemic sclerosis and HIV-1 infection (14 - 16). Siglec-1 can trap HIV-1 particles for trans infection of permissive cells (15).
Varki, A. and T. Angata (2006) Glycobiology 16:1R.
Crocker, P.R. et al. (2007) Nat. Rev. Immunol. 7:255.
Crocker, P.R. et al. (1994) EMBO J. 13:4490.
Hartnell, A. et al. (2001) Blood 97:288.
Nath, D. et al. (1995) J. Biol. Chem. 270:26184.
Crocker, P.R. et al. (1991) EMBO J. 10:1661.
Martinez-Pomares, L. et al. (1999) J. Biol. Chem. 274:35211.
Kumamoto, Y. et al. (2004) J. Biol. Chem. 279:49274.
Nath, D. et al. (1999) Immunology 98:213.
van den Berg, T.K. et al. (2001) J. Immunol. 166:3637.
Nakamura, K. et al. (2002) Glycobiology 12:209.
Barnes, Y.C. et al. (1999) Blood 93:1245.
13. Kirchberger, S. et al. (2005) J. Immunol. 175:1145.
14. York, M.R. et al. (2007) Arthritis Rheum. 56:1010.
15. Rempel, H. et al. (2008) PloS ONE 3:e1967.
16. van der Kuyl, A.C. et al. (2007) Plos ONE 2:e257.
The concentration calculator allows you to quickly calculate the volume, mass or concentration of your vial. Simply enter your mass, volume, or concentration values for your reagent and the calculator will determine the rest.
=
÷
Review this Product
Be the first to review our Recombinant Mouse Siglec-1 Fc Chimera Protein, CF and receive a gift card or discount.













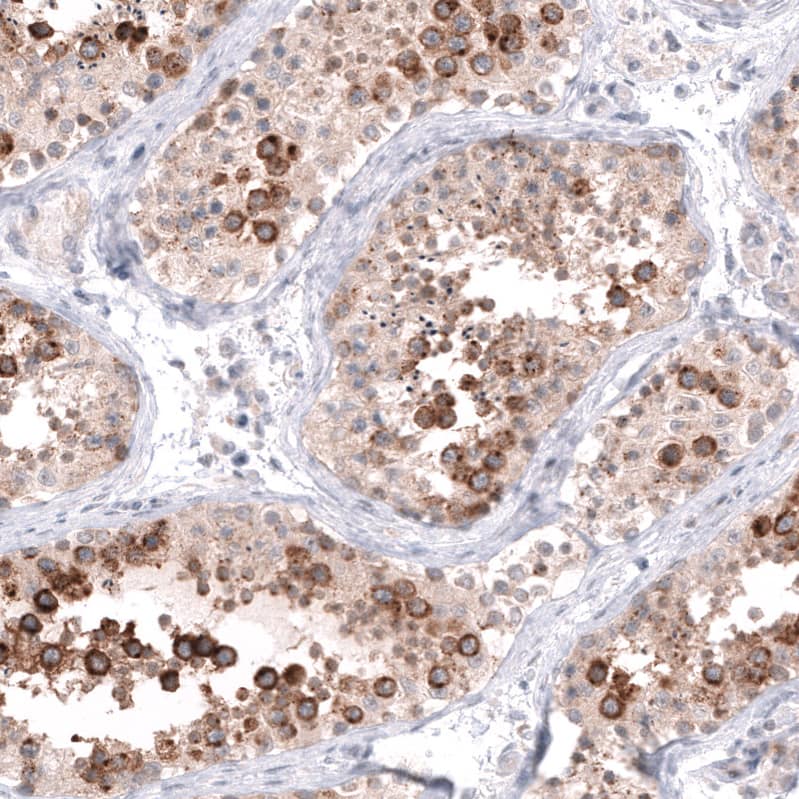
![Bioactivity CTLA-4 [Unconjugated]](https://images.novusbio.com/images/protein/CTLA4_7268CT_2293.jpg)



![Immunocytochemistry CD55/DAF Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/CD55_AF2009_Immunocytochemistry__Immunofluorescence_23052.jpg)
![Immunohistochemistry CD55/DAF Antibody [Unconjugated]](https://images.novusbio.com/images/antibody/CD55_AF2009_Immunohistochemistry_6728.jpg)









![SDS-Page TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNF-alpha_210-TA_256.jpg)
![Bioactivity TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNFalpha_210TA_1658.jpg)
![SEC-MALS TNF-alpha [Unconjugated]](https://images.novusbio.com/images/210-ta_recombinant-human-tnf-alpha-protein-sec-mals-35202312244..jpg)
![Bioactivity IL-2 [Unconjugated]](https://images.novusbio.com/images/202-il_recombinant-human-il-2-protein-bioactivity-174202314946.jpg)
![Immunohistochemistry TFF2 Antibody (366508) [Unconjugated]](https://images.novusbio.com/images/antibody/TFF2_MAB4077_Immunohistochemistry_21485.jpg)

![Flow Cytometry: Siglec-1/CD169 Antibody (HSn 7D2) [NB600-534] - Human CD14+ PBMC differentiated to M1 macrophages with rhGM-CSF were stained with Mouse Anti-Siglec-1/CD169 Monoclonal Antibody (NB600-534, filled histogram), or Mouse IgG1 isotype control (MAB002, open histogram) followed by APC-conjugated Anti-Mouse IgG Secondary Antibody (F0101B).](https://images.novusbio.com/images/Siglec-1-CD169-Antibody-HSn-7D2-Flow-Cytometry-NB600-534-img0002.jpg "Flow Cytometry: Siglec-1/CD169 Antibody (HSn 7D2) [NB600-534] - Human CD14+ PBMC differentiated to M1 macrophages with rhGM-CSF were stained with Mouse Anti-Siglec-1/CD169 Monoclonal Antibody (NB600-534, filled histogram), or Mouse IgG1 isotype control (MAB002, open histogram) followed by APC-conjugated Anti-Mouse IgG Secondary Antibody (F0101B).")