| Reactivity | MuSpecies Glossary |
| Applications | ELISA |
| Conjugate | HRP |
| Background | The Quantikine Mouse IL-1ra Immunoassay is a 4.5 hour solid-phase ELISA designed to measure mouse IL-1ra in cell culture supernates, serum, and plasma. It contains E. coli-expressed recombinant mouse IL-1ra and antibodies raised against the recombinant factor. This immunoassay has been shown to accurately quantitate the recombinant factor. Results obtained using natural mouse IL-1ra... showed linear curves that were parallel to the standard curves obtained using the Quantikine kit standards. These results indicate that this kit can be used to determine relative mass values for naturally occurring mouse IL-1ra. Show More |
| Specificity | Natural and recombinant mouse IL-1ra |
| Source | N/A |
| Assay Type | Solid Phase Sandwich ELISA |
| Inter-Assay | See PDF Datasheet for details |
| Intra-Assay | See PDF Datasheet for details |
| Spike Recovery | See PDF Datasheet for details |
| Sample Volume | See PDF Datasheet for details |
| Gene | Il1rn |
| Dilutions |
|
|
| Application Notes | Interference observed with 1 or more available related molecules. |
|
| Publications |
|
| Storage | Store the unopened product at 2 - 8 °C. Do not use past expiration date. |





| Gene Symbol | Il1rn |




![Bioactivity IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/protein/IL-1_beta_201-LB_306.jpg)
![SDS-Page IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/protein/IL-1_beta_201-LB_304.jpg)
![SEC-MALS IL-1 beta/IL-1F2 [Unconjugated]](https://images.novusbio.com/images/201-lb_recombinant-human-il-1-beta-il-1f2-protein-sec-mals-123202491556.jpg)
![SDS-Page TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNF-alpha_210-TA_256.jpg)
![Bioactivity TNF-alpha [Unconjugated]](https://images.novusbio.com/images/protein/TNFalpha_210TA_1658.jpg)
![SEC-MALS TNF-alpha [Unconjugated]](https://images.novusbio.com/images/210-ta_recombinant-human-tnf-alpha-protein-sec-mals-35202312244..jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/DATA_IL6_M6000_ELISA_936.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/elisa/IL-6_M6000_ELISA_415.jpg)
![N/A IL-6 [HRP]](https://images.novusbio.com/images/m6000b_mouse-il-6-quantikine-elisa-kit-1752025024034.jpg)
![N/A IL-10 [Biotin]](https://images.novusbio.com/images/elisa/DATA_IL10_DY417_ELISA_2014.jpg)
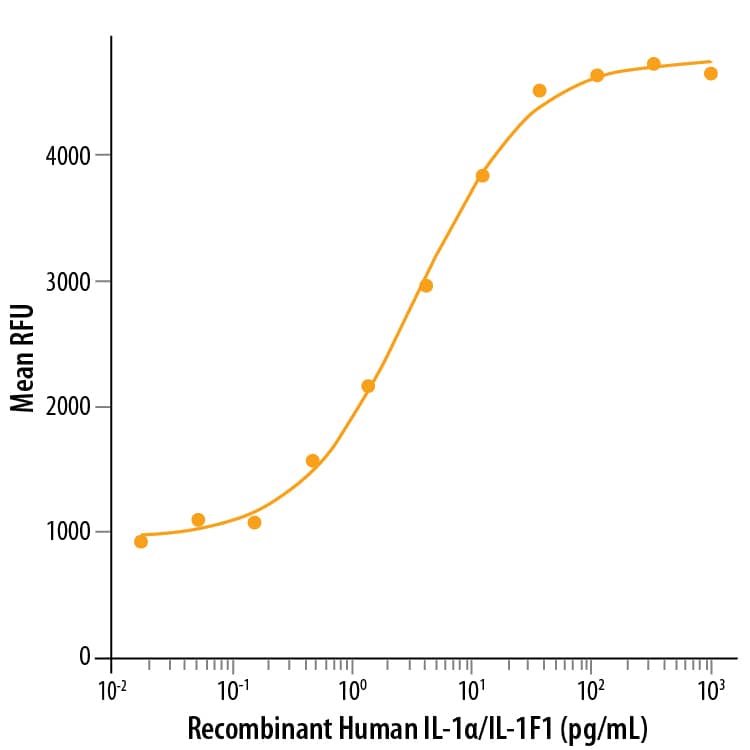
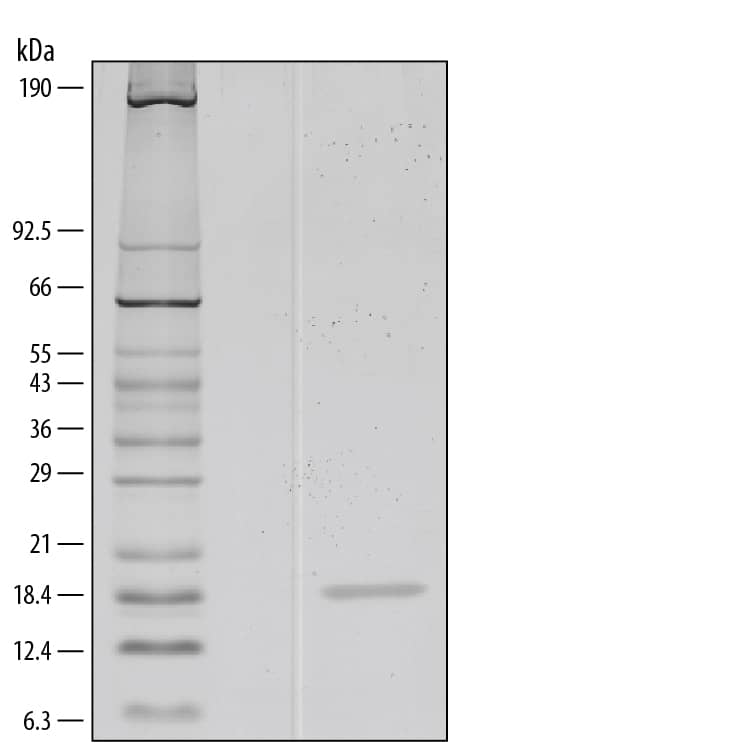
![Bioactivity IL-4 [Unconjugated]](https://images.novusbio.com/images/protein/6507-ilcf_recombinant-human-il-4-cho-expressed-protein-cf-bioactivity-272020133214.jpg)
![Bioactivity IL-2 [Unconjugated]](https://images.novusbio.com/images/202-il_recombinant-human-il-2-protein-bioactivity-174202314946.jpg)
![N/A C-Reactive Protein/CRP [Biotin]](https://images.novusbio.com/images/elisa/DATA_CReactive_Protein_DY1707_ELISA_1701.jpg)
![SEC-MALS IFN-gamma [Unconjugated]](https://images.novusbio.com/images/485-mi_recombinant-mouse-ifn-gamma-protein-sec-mals-1612202583245.jpg)
![Bioactivity IFN-gamma [Unconjugated]](https://images.novusbio.com/images/protein/IFN-gamma_485-MI_455.jpg)
![SDS-Page IFN-gamma [Unconjugated]](https://images.novusbio.com/images/protein/IFN-gamma_485-MI_407.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/CCL2_DCP00_ELISA_67.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/DATA_CCL2_DCP00_ELISA_633.jpg)
![N/A CCL2/JE/MCP-1 [HRP]](https://images.novusbio.com/images/elisa/DATA_CCL2_DCP00_ELISA_634.jpg)
![Bioactivity CTLA-4 [Unconjugated]](https://images.novusbio.com/images/protein/CTLA4_7268CT_2293.jpg)
![Bioactivity GM-CSF [Unconjugated]](https://images.novusbio.com/images/protein/7954-gmcf_recombinant-human-gm-csf-cho-expressed-protein-cf-bioactivity-272020121514.jpg)