 |
By Christina Towers, PhD.
Cells use the complex process of autophagy to degrade and recycle cytoplasmic material. There are over 20 proteins that have been implicated in this process and appropriately named core autophagy genes (ATGs). Most of these were originally discovered in yeast, arguably the most famous being ATG8. In higher eukaryotes ATG8 evolved into a family of proteins known as the GABA type A receptor-associated protein (GABARAP), which includes the Microtubule Associated Protein 1 light chain 3 (MAP1LC3), better known as LC3. Upon starvation or other autophagy inducing stressors, pro-LC3 is cleaved in the cytoplasm by the protease ATG4B creating the LC3-I form which can then be conjugated to phosphatidylethanolamine (PE) by the E1-like enzyme ATG7. The LC3-PE conjugate is thought to be necessary for the canonical formation of autophagosomes. Autophagy is typically defined as the process by which cells degrade cytoplasmic constituent’s and therefore a process confined to the cytoplasm. However, it has been observed by many in the field that a large portion of cellular LC3 resides in the nucleus, raising many questions as to the purpose and function of this nuclear pool of LC3.
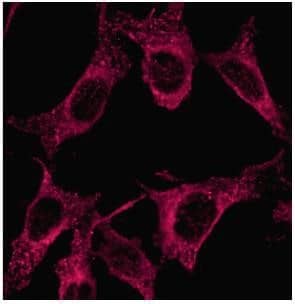 Immunocytochemistry/Immunofluorescence: LC3A Antibody [NB100-2331] - Analysis in PFA fixed NIH/3T3 cells using anti-LC3A antibody.
Immunocytochemistry/Immunofluorescence: LC3A Antibody [NB100-2331] - Analysis in PFA fixed NIH/3T3 cells using anti-LC3A antibody.
In 2015 Huang et al noted that under basal conditions LC3 shuttles between the nucleus and cytoplasm, with apparent even distributions between the two cellular compartments(1). However, under starved conditions LC3 is largely excluded from the nucleus and isolated to the cytoplasm. The authors go on to eloquently show that in nutrient deplete conditions, nuclear LC3 is deacetylated by the nuclear deacetylase, SIRT1 which facilitates its nuclear export. Importantly, these studies show that this nuclear post-translational modification is not only necessary for its nuclear export, but also essential for the LC3-ATG7 interaction: an indispensable interaction for canonical autophagy. These studies therefore suggest that nuclear LC3 is a critical aspect of autophagy.
But with so much LC3 residing in the nucleus, the question still remains, is this nuclear pool of LC3 functional? Around the same time in late 2015, Dou et al started to address this question. They showed that LC3 can indeed function in the nucleus and facilitate the degradation of nuclear lamina components(2). These effects are context dependent and do not occur under basal conditions, but the authors describe these interactions after oncogene activation. These findings have begun to shed light on the functions of this nuclear pool of LC3, and other recent studies are clarifying the mechanisms even further. Studies are now trying to work out the specific residues of LC3 that are important for nuclear localization and transport(3) as well as other nuclear interacting proteins(4). Together these studies conclude that nuclear LC3 is not an artifact but instead an important biologically relevant finding that is involved in canonical autophagy, and likely in other processes as well.
Learn more with Autophagy Handbook
 Christina Towers, PhD
Christina Towers, PhD
University of Colorado (AMC)
Dr. Towers studies the roles of autophagy, apoptosis and cell death in cancer.
References
